I have found enormously entertaining playing with AI to generate a logo for the lab. Not as a straigh forward task as I was hoping initially. So far, my favourite is the one posted below, although it would still need some tweaking… I felt that I lacked the right lexicon in graphic design to be able to express instructions accurately.

Life Anew - A Science Fiction Short Story
Do you like Science Fiction?
Last autumn, I started a part-time evening MA in Creative Writing at University of Birkbeck. It’s been a fantastic experience as writing is something I’ve always enjoyed, both for work and for leisure. I submitted this 4000 word story as my final assignment for my first module in the MA. It got a distinction, so at least my examiners did like it, which I’m pretty happy about!
The story is about two Colombian biologists from an unlikely origin, discovering a second form of life in the Solar System. It happens after a mission to the moon of Jupiter, Europa, brings back some samples for study.
A goal I have for the MA is to explore with my writing alternative Latin American futures, where we take centre stage in the building of knowledge and innovation.
You can read the story by clicking in the link below. If you read it, do let me know what you think, and if you liked it, feel free to share!
Download here:

A photo of Europa by NASA’s Juno Spacecraft (2022)
Evolution of photosynthesis: some brief historic considerations – Part 1
If you are reading this blog post, you might be aware that I have challenged the current accepted view that oxygenic photosynthesis originated from anoxygenic photosynthesis. I have argued, perhaps not yet eloquently enough, that the data on this is clear and straightforward, but the reason it is not widely recognised has nothing to do with the nature of the data, being inconclusive, but it has to do with the history of the subject. The history of how current views on the origin of photosynthesis became first stablished and how these were subsequently modified to accommodate emerging data. Because the idea that “anoxygenic photosynthesis gave rise to oxygenic photosynthesis”, in the sense we understand it today is incorrect [I dare to say with confidence now], the interpretation of new results through flawed frameworks of thought leads—of course—to contradicting and conflicting evolutionary scenarios. Therefore, the subject appears, and has become, controversial and confusing.
I want to give you here a little bit of a taste of how our current views on the evolution of photosynthesis started to become established. I will share with you a text from Cornelis Bernardus van Niel (1897-1985). A key historical figure in photosynthesis research: he basically discovered and described anoxygenic photosynthesis, for example. Not only he was an incredible scientist, but he was also, for what I gather, a highly reputed, celebrated, amazing, and sought after teacher! For what I can understand, he probably taught many of the key scientist who then themselves established “modern photosynthesis research” leading to the second half of the 20th century.
The following is an excerpt from a book chapter written by van Niel and published in 1956. This chapter however is based on things he taught at lectures. The influence of van Niel, either directly or indirectly, is clearly observed in one of the most, if not the most, influential paper on the evolution of photosynthesis to date, by John M. Olson, published in 1970 in Science. Interestingly, Olson dedicated his 1970 paper to van Niel. Olson, then became a big influence on Prof. Robert Blankenship, who wrote what I consider to be the second most influential paper on the subject, published in 1992 in Photosynthesis research. While Blankenship and Olson did not necessarily agree on all the details, the overall framework of thinking in the 1992 paper, is indeed somewhat derived from Olson and colleagues’ writings starting with that 1970 paper. Van Niel himself was also very influenced by the ideas of other scientists of his time. For example, by Norman H. Horowitz, as in the “Horowitz hypothesis” of biochemical evolution, which van Niel acknowledged as correct in the quoted chapter below.
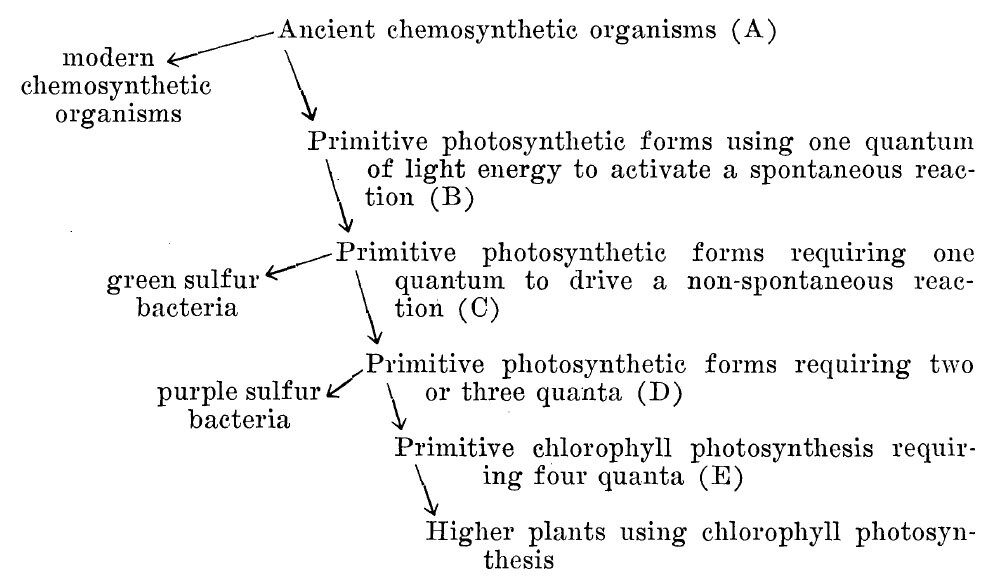
Early concepts on the evolution of photosynthesis, by Harold F. Blum, 1937, published in The American Naturalist, 71:350-362. This scheme was produced using data from van Niel’s experiments characterizing anoxygenic photosynthesis!
From “Chapter 6: Evolution as viewed by the microbiologist” by C. B. van Niel, 1956. In The Microbe's Contribution to Biology. Harvard University Press. A part of John M. Prather Lecture Series in Biology.
Now, the newer concepts are important partly because they make the earlier failures comprehensible, partly also because they are in line with present-day knowledge of biochemical events. In this respect they contrast favorably with previous attempts which, based on an acceptance of biopoesis*, were concerned with speculations on the probable nature of the first living organisms on earth. The starting point for these speculations used to be the idea that the earth was a purely mineral environment at the time when it had sufficiently cooled to sustain life. Such an environment is, of course, unsuitable for the growth of any but the chemo- and photoautotrophic organisms. This led to the conclusion that microbial representatives of these groups had to be considered as the most likely progenitors of all other forms of life. Only after the former had produced organic matter from the inorganic constituents of the earth's atmosphere and lithosphère was an environment created in which organisms with requirements for organic substances could grow.
Ostensibly this sort of reasoning satisfies the demands of the evolutionary theory in which the more complex is derived from the simpler. Nevertheless, there is a serious flaw in the argument, and this has become more and more obvious as our understanding of biochemical reaction mechanisms has increased. We have learned to interpret these mechanisms as chains of step reactions, each one controlled by a specific catalyst or enzyme, and we have come to think of differences in nutritional requirements in terms of differences in synthetic ability of the organisms concerned. The evidence provided by studies with a wide variety of microbes even suggests that the greater is the biosynthetic capacity of an organism, the more elaborate must be its enzymatic composition. From this it follows that the chemo- and photoautotrophs, which exhibit the greatest biosynthetic ability known to us at present, must possess a vast array of enzymes. Once this is granted, it is also clear that such an organism cannot be considered as "primitive," and that we must qualify as wholly inadequate the concept that the first living creatures were of this kind. The evolutionary approach to biopoesis dictated the need for starting with a simple system. But the above analysis shows that the selection of the environment for this purpose entails the inevitable consequence that the first organism must have been exceedingly complex.
It is probable that aerobic metabolism did not evolve until green-plant photosynthesis had created an environment in which oxygen was available. The first biochemical patterns would thus be representative of the diverse types of anaerobic processes. In line with our primary assumption, we must grant that entities with the most complex nutrient requirements, that is, with the least developed biosynthetic activity and the greatest dependence on a complex environment, antedated those with more restricted needs. The first phase of biochemical evolution would therefore have involved the gradual acquisition of the ability to use progressively less complex mixtures of organic ingredients for the production of the chemical constituents of the organisms. This phase can thus be characterized as an evolution toward increased independence on a medium replete with a multitude of organic materials. The theoretical limit of such independence is reached with the appearance of organisms that no longer require any organic substances. These are the chemo- and photoautotrophic organisms. Among the anaerobic microbes known at present this property is exhibited by the causative agents of sulfate reduction and of the methane fermentation, by the green and purple sulfur bacteria, and by the algae. Far from being "primitive," the metabolic characteristics of such organisms now appear to occupy a position near the end of the scale of biochemical evolution.
*Biopoesis: "the creation of something, that some people might wish to call living, from non-living material.” [As defined in the chapter].
What happens then is that Olson fits whatever little was known in 1970 about photosystem function, and it was very very very little, into a framework of thought that is nearly identical to van Niel’s, but I’m not sure is entirely attributable to van Niel himself, perhaps it was more like an amalgamation of ideas being discussed at the time. Since then, every discussion on photosystem evolution is fitted into Olson’s old framework, one way or another, changing one group’s position or a photosystem’s position in the scheme, but overall the same conceptual framework… until yours truly enters the scene to reassess the topic beginning with my Photosynthesis research paper published in 2015. However, I was a very immature writer at the time, and I still am... for sure! Meaning that the good points I made, kind of got diluted amid other less relevant stuff. In any case, my work is basically about slowly breaking away from this framework. Or at least, that is what I want to believe.
Olson in his 1970’s paper introduced the idea of photosystem evolution for the first time. He proposed that “system 1” gave rise to “system 2”. System 1 referring to what we now know as type I reaction centres, although that was not entirely well defined then. “System 2” referred to water-splitting photosystem II, because then it was not really known that purple bacteria had an anoxygenic type II reaction centre that was homologous to photosystem II. Then once anoxygenic type II reaction centres were described properly later, it was easy to fit into its evolutionary position between “system 1” and the water-splitting “system 2”, as it’s evident in the current standard thinking on the evolution of photosynthesis.

An evolutionary scenario. This is by Pierson and Olson, in a book chapter from 1989, but this one is elaborated on his 1970 ideas. The same figure was reprinted in 1992’s Blankenship paper.
As you can see now, these ideas do not emerge from a proper evolutionary treatment of data. What is more, none of these early speculations on the evolution of photosynthesis were really led by evolutionary biologists. It means that the researchers unconsciously leant towards describing evolution as a linear series of events, which is often not appropriate. The first phylogeny of type II reaction centres was not published until 1990 and was well interpreted but had little impact on the literature or evolutionary thinking, unlike Blankenship’s 1992 paper. A more appropriate and detailed interpretation of this did not happen until my own work… in fact, I have spent nearly all of my independent career as a scientists, about eight years now, studying just that one phylogeny.
To be continued…

The synthesis of my work published in BBA -Bionergetics, 2021.